|
|
| Miembros del equipo: Nieves Fernández, José Fernández , Cristina Martón, Carmen Herrero, Sara Alonso, Alicia Azogil, Saioa Márquez y Mariano Sánchez Crespo. |

- i) La descripción del efecto modulador negativo de la unión covalente del factor 3 del complemento a la porción Fc de la IgG sobre la capacidad de los macrófagos humanos para liberar mediadores inflamatorios. Este hecho se explica por la derivación de la carga de complejos inmunes del receptor FcyR al receptor 3 del complemento (CR3, αMβ2 integrina), cuya capacidad para captar partículas por fagocitosis se asocia con una débil inducción de la liberación de mediadores inflamatorios.
- ii) La potenciación de los efectos proinflamatorios de los β-glucanos por componentes del sistema del complemento mediante la cooperación entre el receptor de β-glucanos dectin-1 y CR3.
- iii) La activación de la transcripción dependiente de C/EBPβ y NF-kB por el cross-linking de los receptors para la porción Fc de la molécula de IgG y su asociación con la regulación transcripcional de las quemocinas MIP-1α/CCL3, MIP-1β/CCL4, RANTES/CCL5 e IL-8/CXCL8 por las células monocíticas.
- iv) La descripción de la limitada capacidad de los lipopolisacáridos de Brucella spp. y de lipopolisacáridos con estructuras similares de su lípido A para activar los receptores TLR-4 (Toll-like receptor-4).
- v) La descripción de nuevos mecanismos de regulación del metabolismo del ácido araquidónico en los leucocitos polimorfonucleares humano por ligandos del sistema inmune innato como el peptidoglicano y los patrones que incluyen restos de manosa.
|
|
|
Estructuras químicas de los lípidos A. Las estructuras correspondientes a los lípidos A de los lipopolisacáridos de Escherichia coli, Brucella/Ochrobactrum y Francisella tularensis se muestran de acuerdo con los datos publicados por Zhou et al., Moriyon, Velasco et al., y Vinogradov et al. Las principales diferencias estructurales se refieren a la presencia de grupos fosfato, al esqueleto de disacáridos y al número y longitud de las cadenas de ácidos grasos. |
| Distribución del marcaje con [3H]AA en las distintas fracciones lipídicas de los polimorfonucleares y en los sobrenadantes . Los PMN marcados con [3H]AA se estimularon durante una hora en presencia de 10 μg/ml de PGN y con 25 mg/ml de mannan, o se mantuvieron en medio. Al cabo de este tiempo, los pellets celulares se separaron por centrifugación. Los extractos lipídicos de los pellets y de los sobrenadants se usaron para separación por cromatografía en capa fina. La migración de los patrones en la capa fina se muestra en (A). Los polimorfonucleares se incubaron con calpeptin y pyrrolidina-1 antes de la adición de los estímulos y se cuantificó el [3H]AA liberado en los sobrenadantes. Los datos representan media ± ESM de tres experimentos en duplicado *, p < 0.05 (B). El cromatograma de un sobrenadante de polimorfonucleares humanos estimulados con mannan muestra los tiempos de retención de PGE2/PGD2 y 20-OH-LTB4. Ambos compuestos han sido identificados por sus espectros de masa (C). Espectros de fragmentación en el modelo de ion negativo usando MRM (multiple reaction monitoring) de las transiciones específicas m/z 335—195 para LTB4, m/z 351—195 para 20-OH-LTB4, m/z 303—205 para ácido araquidónico, m/z 351—189 para PGE2/PGD2 (D). |
- vi) El mecanismo de inducción de la proteína ciclooxigenasa-2 (COX-2) en polimorfonucleares humanos por los patrones moleculares asociados a patógenos. En este sistema, el peptidoglicano y el mannan, y en menor escala el homólogo de lipoproteínas bacterianas Pam3CSK4, indujeron la expresión de la proteína COX-2. Por el contrario, el ácido lipoteicoico y el muramildipeptido fueron estímulos irrelevantes. El mRNA que codifica la COX-2 estaba presente en los polimorfonucleares en estado basal en niveles similares a los detectados en células estimuladas, mientras que la detección de la proteína era indetectable. El tratamiento con el inhibidor de la fosfatidilinositol-3-kinasa (PI3K) wortmaninn, el inhibidor de mTOR (mammalian target of rapamycin) rapamycin y el inhibidor de la traducción cycloheximida bloqueó la inducción de la proteina COX-2 en respuesta a mannan y peptidoglicano, mientras que el inhibidor de la transcripción actinomycin D no mostró ningún efecto. Estos resultados han puesto de manifiesto la capacidad de los patrones moleculares asociados a patógenos para inducir el metabolismo oxidativo del ácido araquidónico de forma más robusta que los quimiotrayentes clásicos de polimorfonucleares, puesto que el mannan y el peptidoglicano hacen coincidir la liberación de ácido araquidónico con la rápida inducción de la proteína COX-2, regulada por una cascada de señalización que incluye PI3K, mTOR, y la maquinaria de traducción. Este mecanismo de inducción de la expresión de la proteína COX-2 en PMN es sustancialmente diferente del que se produce en fagocitos mononucleares, que depende, fundamentalmente, de la regulación transcripcional.
- vii) La caracterización de los mecanismos moleculares que explican las distintas respuestas de las células dendríticas derivadas de monocitos a los patrones moleculares derivados de hongos que contienen manosa y polímeros de β-glucanos. Esta respuesta se caracteriza por la prominente inducción de COX-2, la alta producción de IL-10 e IL-23 y una reducida biosíntesis de IL-12 p70. La unión cooperativa de dectin-1 por β-glucanos y de DC-SIGN por le componente de manosa parece crucial para la inducción de estas respuestas.
| Agrupamiento de receptores que reconocen el zymosan en células dendríticas. Las células se incubaron con zymosan marcado con Alexa-Fluor® 488 (verde) y se tiñeron con mAb frente a anti-DC-SIGN, anti-dectin-1 y anti-receptor de manosa y, posteriormente con anticuerpo de cabra anti-ratón marcado con Alexa-Fluor® 594 (rojo). Tras la estimulación por zymosan, todos los receptores se agrupan inicialmente alrededor de la partícula. Posteriormente, la tinción de DC-SIGN no se agrupa alrededor del zymosan (puntas de flecha blancas), lo que indicaría que aunque interviene en el reconocimiento de la partícula, no es necesario para la captación. |
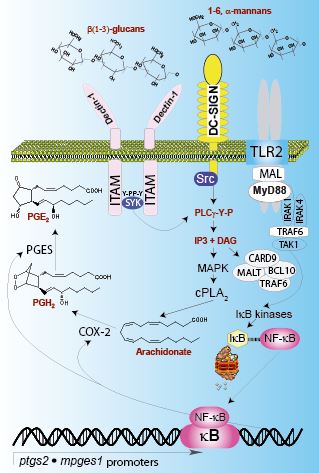 The Candida paradigm for AA metabolism in DC. The mannan and β-glucan components of the fungal cell wall are recognized by at least DC-SIGN, TLR2, dectin-1, the mannose receptor, and dectin-2. This gives rise to a series of signalling events implicating activation of SYK and Src families of tyrosine kinases. Both routes converge to activate phospholipases Cγ and through the generation of diacylglycerol activate protein kinase C and MAPK cascades. Phosphorylation by MAPK and Ca2+-driven translocation of cPLA2α explain AA release from cell phospholipids. Activation of IκB kinases via MyD88 is a major factor explaining COX-2 and PGES induction. The CARD9/MALT/BCL10/TRAF6 complex is also involved in the activation of IκB kinases. DAG, diacylglycerol; IP3, inositol 1,4,5-trisphosphate; IκB, inhibitory subunit of NF-κB; IRAK, interleukin-1 receptor-associated kinase; MAL, MyD88 adaptor-like; MAPK, mitogen-activated protein kinases; MyD88, myeloid differentiation primary response gene 88; PLCγ, phospholipase Cγ; P-Y, phosphotyrosine; ptgs2, COX-2 gene; mpges1, microsomal prostaglandin E synthase-1 gene; TAK1, transforming growth factor-β-activated kinase-1; TRAF6, TNF receptor-associated factor-6. This scheme has been constructed from the results reported by Valera et al. (J Immunol180: 5727-5736. 2008), Suram et al. (J Biol Chem281: 5506-5514, 2006 & J Biol Chem285: 30676-30685, 2010), and Parti et al. (Am J Respir Crit Care Med42: 415-423, 2011).
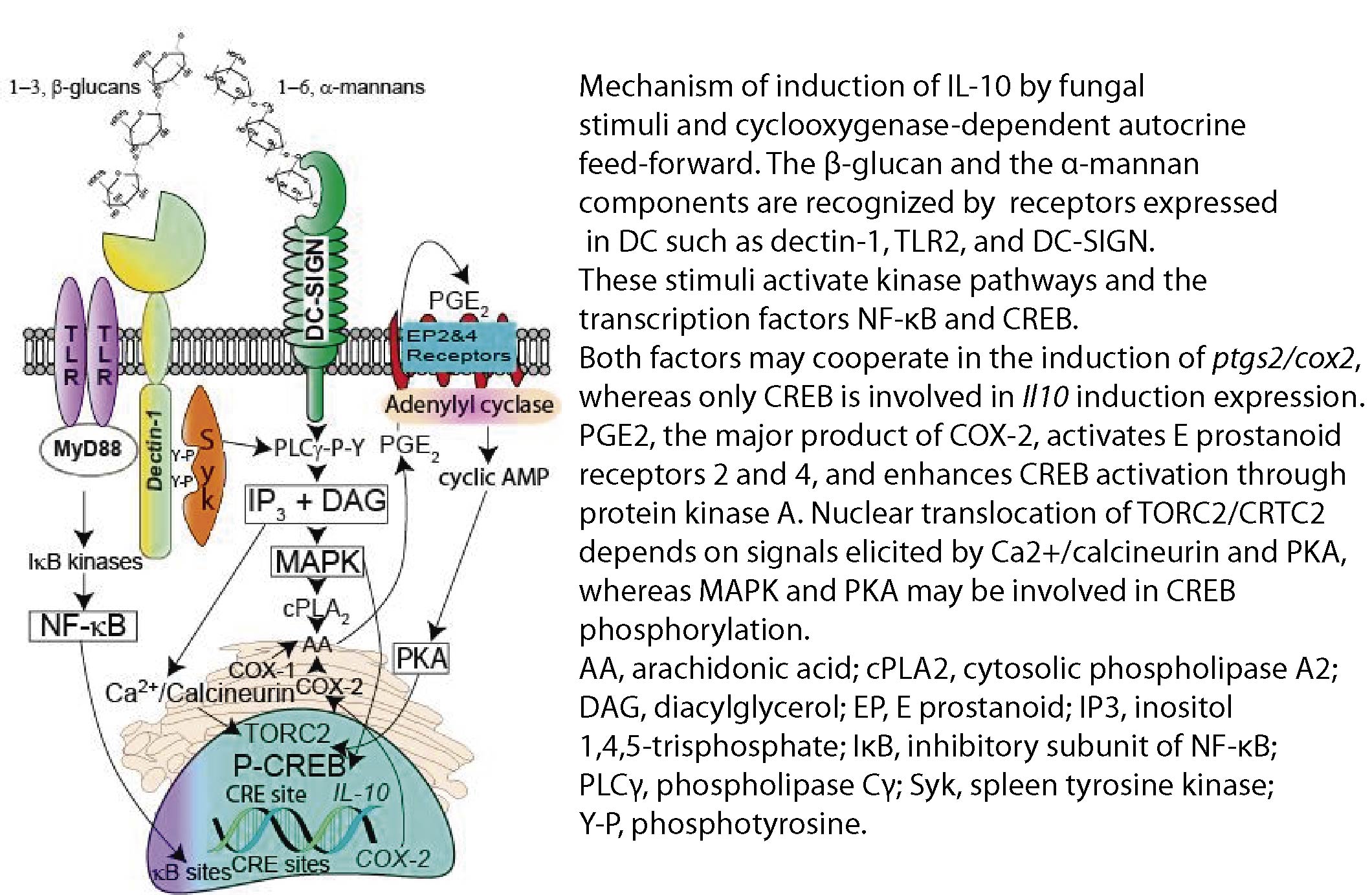
La alta producción de IL-10 asociada a la baja producción de IL-12 p70 puede explicarse por un mecanismo que implica la secreción autocrina de PGE2 y una red de interacciones entre PKA, MAP/ERK, (GSK3β) glicógeno sintasa kinasa-3β y calcineurina que regulan la actividad transcripcional de CREB (CRE binding protein) mediante su capacidad para unir los coactivadores CBP (CREB-binding protein) y CRTC2 (CREB regulated transcription coactivator, también denominado TORC2, transducer of regulated CREB activity) y disminuir la cantidad de CBP disponible para la interacción con otros factores de transcripción.
The Candida paradigm for AA metabolism in DC. The mannan and β-glucan components of the fungal cell wall are recognized by at least DC-SIGN, TLR2, dectin-1, the mannose receptor, and dectin-2. This gives rise to a series of signalling events implicating activation of SYK and Src families of tyrosine kinases. Both routes converge to activate phospholipases Cγ and through the generation of diacylglycerol activate protein kinase C and MAPK cascades. Phosphorylation by MAPK and Ca2+-driven translocation of cPLA2α explain AA release from cell phospholipids. Activation of IκB kinases via MyD88 is a major factor explaining COX-2 and PGES induction. The CARD9/MALT/BCL10/TRAF6 complex is also involved in the activation of IκB kinases. DAG, diacylglycerol; IP3, inositol 1,4,5-trisphosphate; IκB, inhibitory subunit of NF-κB; IRAK, interleukin-1 receptor-associated kinase; MAL, MyD88 adaptor-like; MAPK, mitogen-activated protein kinases; MyD88, myeloid differentiation primary response gene 88; PLCγ, phospholipase Cγ; P-Y, phosphotyrosine; ptgs2, COX-2 gene; mpges1, microsomal prostaglandin E synthase-1 gene; TAK1, transforming growth factor-β-activated kinase-1; TRAF6, TNF receptor-associated factor-6. This scheme has been constructed from the results reported by Valera et al. (J Immunol180: 5727-5736. 2008), Suram et al. (J Biol Chem281: 5506-5514, 2006 & J Biol Chem285: 30676-30685, 2010), and Parti et al. (Am J Respir Crit Care Med42: 415-423, 2011).
La alta producción de IL-10 asociada a la baja producción de IL-12 p70 puede explicarse por un mecanismo que implica la secreción autocrina de PGE2 y una red de interacciones entre PKA, MAP/ERK, (GSK3β) glicógeno sintasa kinasa-3β y calcineurina que regulan la actividad transcripcional de CREB (CRE binding protein) mediante su capacidad para unir los coactivadores CBP (CREB-binding protein) y CRTC2 (CREB regulated transcription coactivator, también denominado TORC2, transducer of regulated CREB activity) y disminuir la cantidad de CBP disponible para la interacción con otros factores de transcripción.
 La investigación actual se dirige al análisis de las vías de señalización acopladas a la activación de los receptores del sistema inmune innato expresados en la superficie de las células dendríticas y a la caracterización de los mecanismos de regulación de la producción de citocinas que influyen en al polarización de la respuesta inmune hacia los tipos Th1, Th2 y Th17. En este sentido, recientemente hemos definido:
La investigación actual se dirige al análisis de las vías de señalización acopladas a la activación de los receptores del sistema inmune innato expresados en la superficie de las células dendríticas y a la caracterización de los mecanismos de regulación de la producción de citocinas que influyen en al polarización de la respuesta inmune hacia los tipos Th1, Th2 y Th17. En este sentido, recientemente hemos definido:
- El mecanismo de regulación transcripcional de IL23A (gen que codifica la cadena p19 de la citocina IL-23) mediante fosforilaciones complementarias del factor ATF.
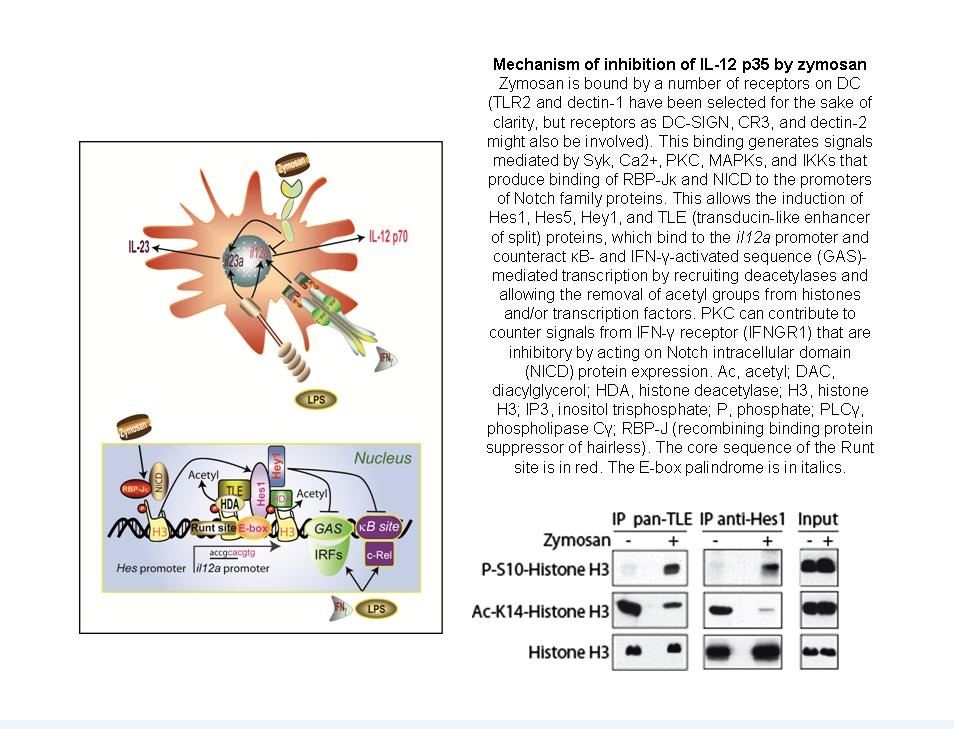
 2. La activación de SIRT1 durante la fagocitosis de patrones moleculares de la pared de hongos y su contribución a la represión de la transcripción de la cadena p35 de la citocina IL-12 p70.
2. La activación de SIRT1 durante la fagocitosis de patrones moleculares de la pared de hongos y su contribución a la represión de la transcripción de la cadena p35 de la citocina IL-12 p70.
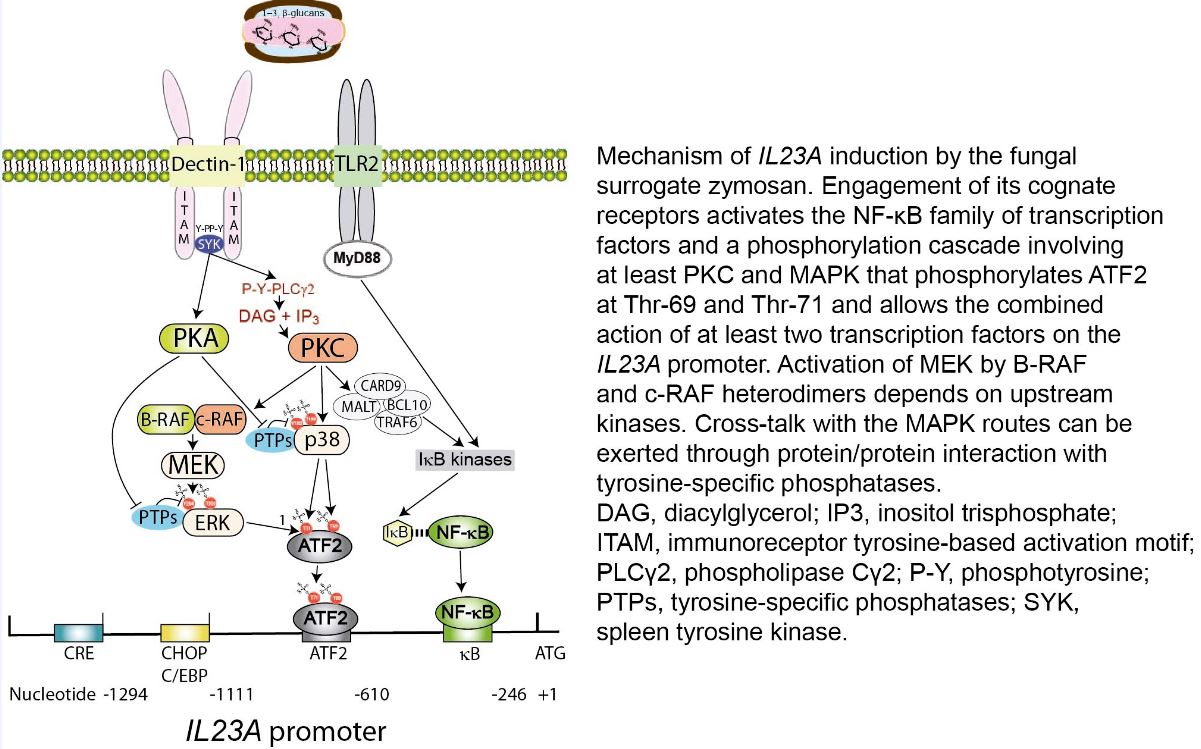
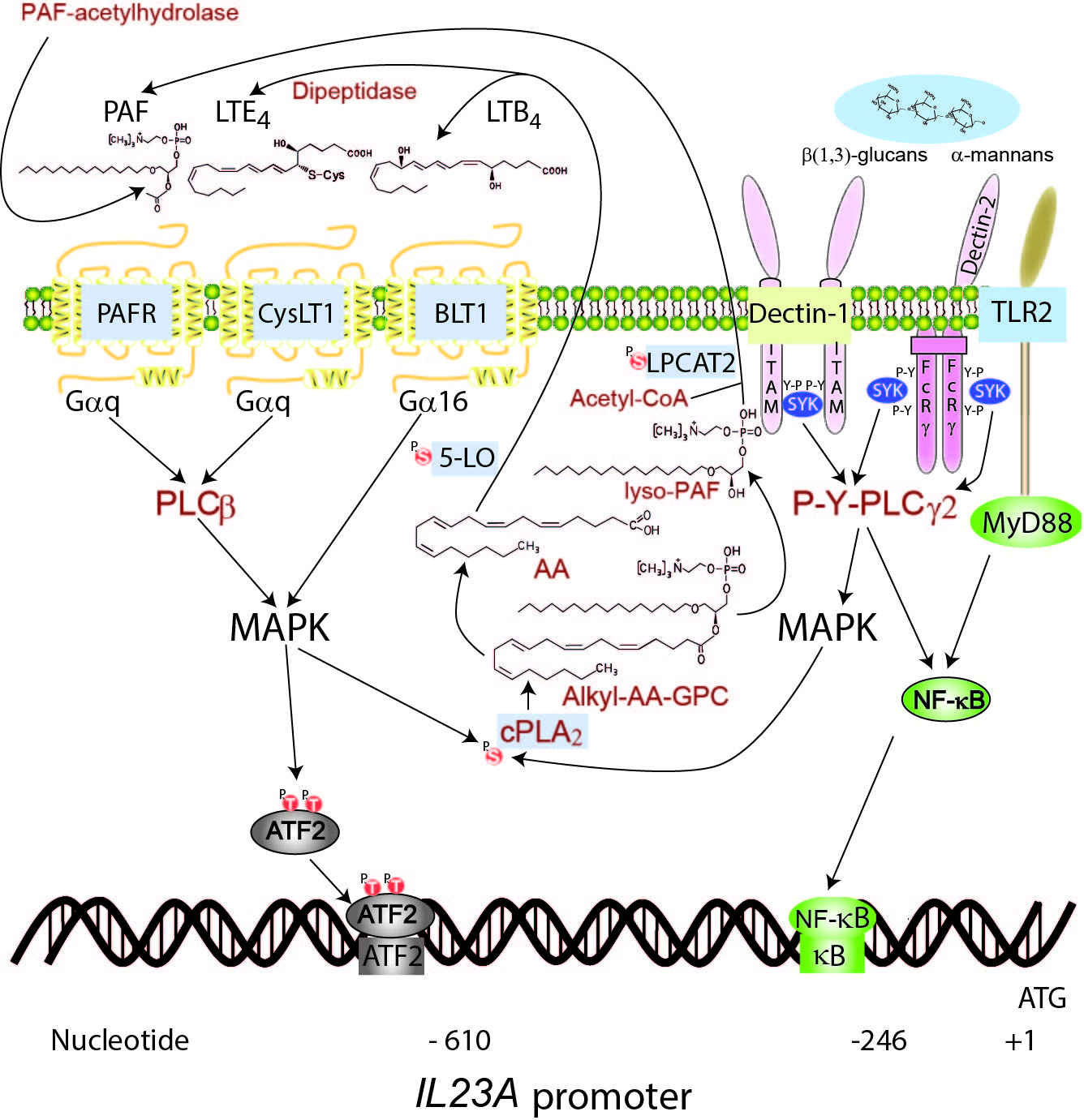 Cooperation of autocrine lipid mediators with fungal patterns to induce ATF2 phosphorylation and the trans-activation of IL23A through complementary phosphorylations of activating transcription factor 2 (ATF2). Fungal patterns activate the NF-κB family of transcription factors and a phosphorylation cascade involving at least PKC and MAPK that phosphorylates ATF2 at Thr69 and Thr71, cPLA2 at Ser505, 5-LO at Ser663, and LPCAT at Ser34. P-Ser505-cPLA2 hydrolyses 1-hexadecyl-2-arachidonoyl-sn-glycero-3-phosphocholine (hexadecyl-AA-GPC) and generates both free AA and 1-hexadecyl-2-lyso-glycero-3-phosphocholine (lyso-PAF) that by the action of 5-LO and LPCAT1/2 give rise to LTB4, cysteinyl-LT, and PAF. The lipid mediators activate another signaling cascade that further enhances their biosynthesis and the phosphorylation of ATF2. The combined action of NF-kB and ATF2 on the IL23A promoter allows its trans-activation. FcRγ, Fc receptor γ-chain; ITAM, immunoreceptor tyrosine-based activation motif; PLCβ, phospholipase Cβ; PLCγ2, phospholipase Cγ2; P, phosphate; P-Y, phosphotyrosine; SYK, spleen tyrosine kinase; TLR2, Toll-like receptor 2.
Cooperation of autocrine lipid mediators with fungal patterns to induce ATF2 phosphorylation and the trans-activation of IL23A through complementary phosphorylations of activating transcription factor 2 (ATF2). Fungal patterns activate the NF-κB family of transcription factors and a phosphorylation cascade involving at least PKC and MAPK that phosphorylates ATF2 at Thr69 and Thr71, cPLA2 at Ser505, 5-LO at Ser663, and LPCAT at Ser34. P-Ser505-cPLA2 hydrolyses 1-hexadecyl-2-arachidonoyl-sn-glycero-3-phosphocholine (hexadecyl-AA-GPC) and generates both free AA and 1-hexadecyl-2-lyso-glycero-3-phosphocholine (lyso-PAF) that by the action of 5-LO and LPCAT1/2 give rise to LTB4, cysteinyl-LT, and PAF. The lipid mediators activate another signaling cascade that further enhances their biosynthesis and the phosphorylation of ATF2. The combined action of NF-kB and ATF2 on the IL23A promoter allows its trans-activation. FcRγ, Fc receptor γ-chain; ITAM, immunoreceptor tyrosine-based activation motif; PLCβ, phospholipase Cβ; PLCγ2, phospholipase Cγ2; P, phosphate; P-Y, phosphotyrosine; SYK, spleen tyrosine kinase; TLR2, Toll-like receptor 2.
The IL-10-dependent and -independent cross-inhibitory networks triggered by fungal patterns.
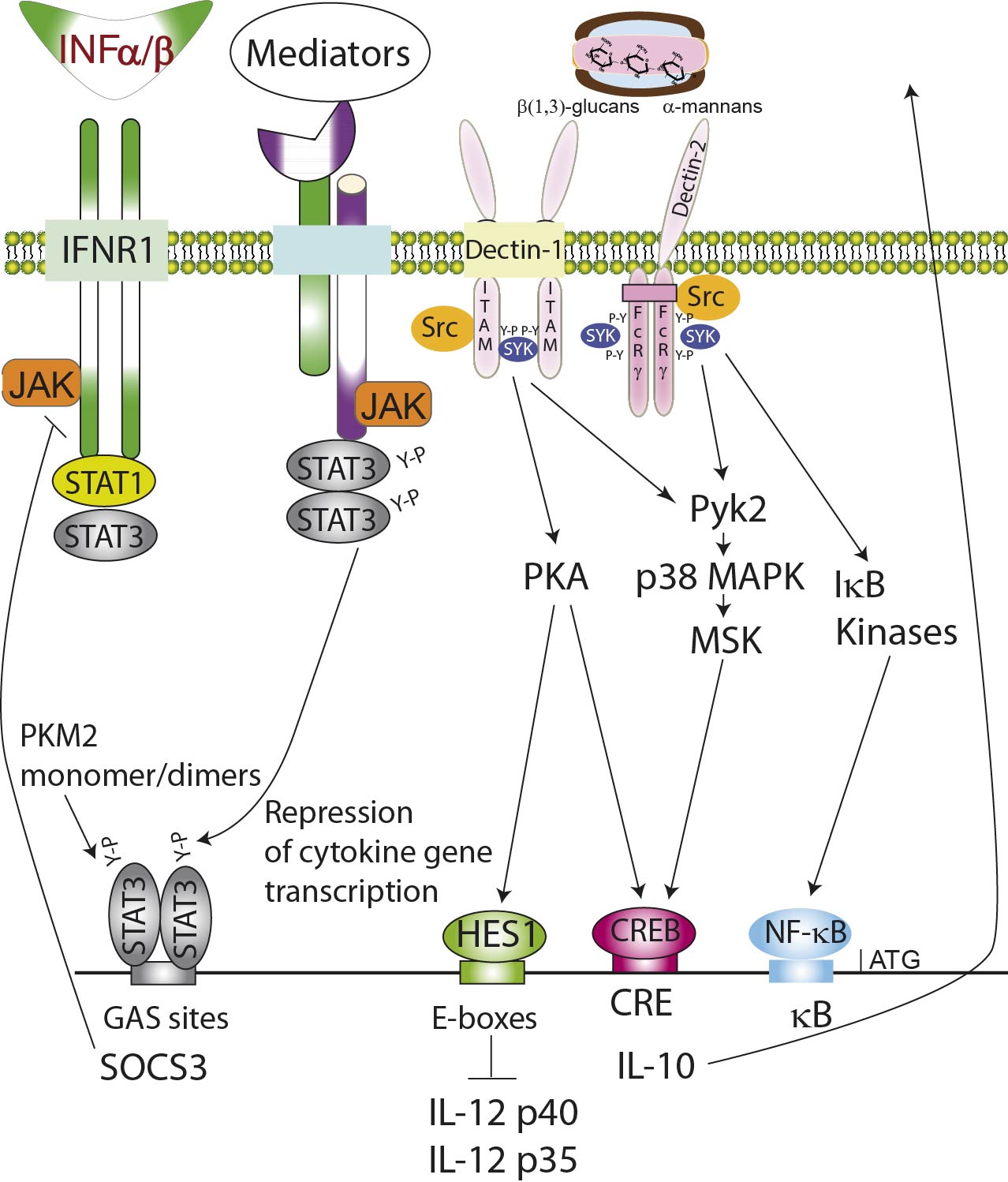
Binding of α-mannan and β-glucan moieties initiates a series of phosphorylation reactions involving MAP kinases, protein kinase A, IkB kinases, as well as nuclear monomer/dimers of the key metabolic regulator pyruvate kinase M2 (PKM2).1 The activation of the latent cytoplasmic transcription factors CREB and NF-kB trans-activates the IL-10 and IL-12 family cytokine promoters. The concomitant activation of repressors of the NOTCH family, for instance HES1,2-4 contributes to blunt the production of IL-12 p70 protein. A network of secondary mediators activates the formation of Y705-STAT3 dimers that both directly and through the transcriptional activation of SOCS genes contribute to the repression of cytokine gene transcription.
References- Gao X, Wang H, Yang JJ, Liu X, Liu ZR. Pyruvate kinase M2 regulates gene transcription by acting as a protein kinase. Mol Cell 2012; 45:598-609.
- Wang L, Gordon RA, Huynh L, Su X, Park Min KH, Han J et al. Indirect inhibition of Toll-like receptor and type I interferon responses by ITAM-coupled receptors and integrins. Immunity 2013; 32:518-30.
- Alvarez Y, Municio C, Hugo E, Zhu J, Alonso S, Hu X et al. Notch- and transducin-like enhancer of split (TLE)-dependent histone deacetylation explain interleukin 12 (IL-12) p70 inhibition by zymosan. J Biol Chem 2011; 286:16583-195.
- Larabee JL, Shakir S., Barua, S, Ballard JD. Increased cAMP in monocytes augments Notch signaling mechanisms by elevating RBP-J and transducin-like enhancer of Split (TLE). J Biol Chem 2013; 288:21526-36.
 El objetivo final de nuestra investigación es la caracterización de los receptores celulares y de las vías de señalización que pueden constituir dianas farmacológicas relevantes para el tratamiento de enfermedades autoinmunes, glomerulonefritis, vasculitis y enfermedades pulmonares de base inflamatoria.
El objetivo final de nuestra investigación es la caracterización de los receptores celulares y de las vías de señalización que pueden constituir dianas farmacológicas relevantes para el tratamiento de enfermedades autoinmunes, glomerulonefritis, vasculitis y enfermedades pulmonares de base inflamatoria.
PUBLICACIONES RECIENTES
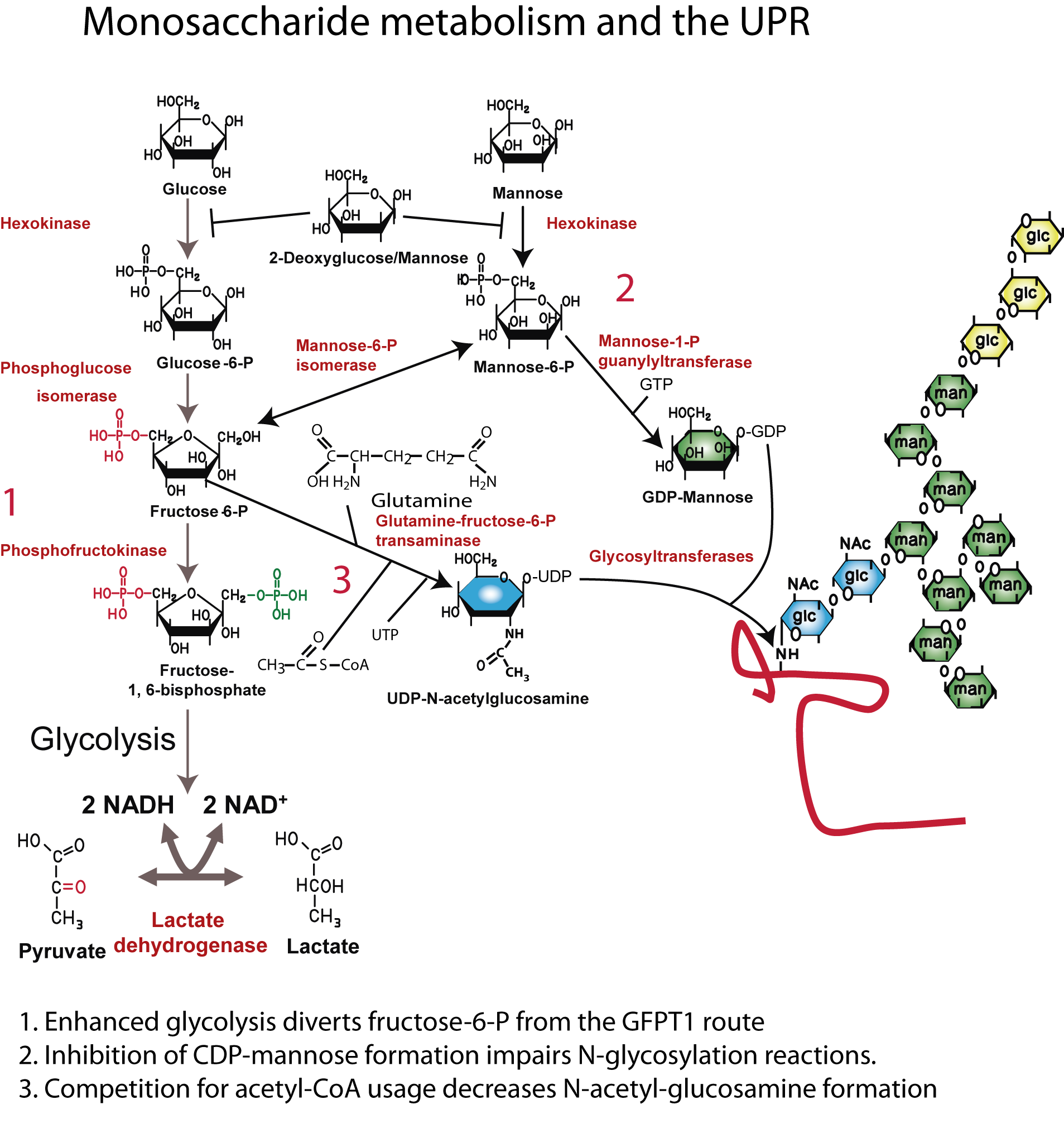
- Marquez, S., Fernández, J.J., Terán-Cabanillas, E., Herrero, C., Alonso, S., Azogil, A., Montero, O., Cubillos-Ruiz, J., Fernández, N., Sánchez Crespo, M. (2017) Endoplasmic reticulum stress sensor IRE1α enhances IL-23 expression by human dendritic cells. Front. Immunol. doi: 10.3389/fimmu.2017.00639.
- Rodríguez, M., Márquez, S. Montero, O., Alonso, S., García Frade, J., Sánchez Crespo, M., Fernández, N. (2016) Pharmacological inhibition of eicosanoids and platelet-activating factor signaling impairs zymosan induced release of IL-23 by dendritic cells. Biochem. Pharmacol. 102:78-96.
- Rodríguez, M, Domingo, E., Alonso, S, Frade, J.G., Eiros, J., Sánchez Crespo, M., Fernández, N. (2014) The unfolded protein response and the phosphorylations of activating transcription factor 2 in the trans-activation of IL23A promoter produced by β-glucans. J. Biol. Chem. 289:22942-22957.
- Rodríguez, M., Domingo, E., Municio, C., Alvarez, Y., Hugo, E., Fernández, N., Sánchez Crespo, M. (2014) Polarization of the innate immune response by prostaglandin E2: a puzzle of receptors and signals. Mol. Pharmacol. 85:187-197.
- Municio, C., Alvarez Y, Montero, O., Hugo, E., Rodríguez, M., Domingo, E., Alonso, S., Fernández, N., Sánchez Crespo, M. (2013) The response of human macrophages to β-glucans depends on the inflammatory milieu. PLoS One 2013:8(4): e62016. doi:10.1371/journal.pone.0062016.
- Alvarez Y, Rodríguez M, Municio C, Hugo E, Alonso S, Ibarrola N, Fernández N, Sánchez Crespo M. (2012) Sirtuin 1 is a key regulator of the IL-12 p70/IL-23 balance in human dendritic cells. J. Biol. Chem. 287:35689-35701.
- Municio, C., Hugo, E., Alvarez, Y., Alonso, S., Blanco, L., Fernández, N., Sánchez Crespo, M. (2011) Apoptotic cells enhance IL-10 and reduce IL-23 production in human dendritic cells treated with zymosan. Mol. Immunol. 49, 97-106.
- Alvarez Y, Municio C, Hugo E, Zhu J, Alonso S, Hu X, Fernández N, Sánchez Crespo M. (2011) Notch and transducin-like enhancer of split (TLE)-dependent histone deacetylation explain IL-12 p70 inhibition by zymosan. J. Biol. Chem. 286, 16583-16595
- Alvarez, Y., Valera, I., Municio, C., Hugo, E., Padrón, F., Blanco, L., Rodríguez, M., Fernández, N., Sánchez Crespo, M. (2010) Eicosanoids in the innate immune response: TLR and non-TLR routes. Mediators Inflamm. doi: 10.1155/2010/201929.